








BEMutatkozás
A Szelenofehérje Kutató Osztály 2020-as megalapítása óta onkológiához kapcsolódó alapkutatásokkal foglalkozik. Osztályvezetőnk Prof. Dr. Elias Arnér svéd kutatóorvos, a szelenofehérjék nemzetközi szaktekintélye.
Prof. Arnér az Országos Onkológiai Intézet felkérésére jelenleg részmunkaidőben irányítja a Szelenofehérje Kutató Osztály munkáját, egyidejűleg a stockholmi Karolinska Intézet és Karolinska Kórház közös rákcentrumának (CRKI) tudományos igazgatója és a Karolinska Intézetben működő másik kutatócsoportjának vezetője is. Az Országos Onkológiai Intézetben jelenleg 5 fős csoport dolgozik – projektjeink szervesen kapcsolódnak Elias Arnér professzor svédországi tevékenységéhez, a stockholmi testvércsoportunkkal rendszeres online és személyes megbeszélésekkel biztosítjuk a folyamatos tudásmegosztást.

Kutatási témáink
Vizsgálataink középpontjában a fehérjék egy speciális csoportja, a szelenofehérjék állnak. Kutatásaink célja ezeknek a fehérjéknek egészséges és patológiás (különösen a daganatos) folyamatokban betöltött szerepének felderítése.
A szelenofehérjék közös jellemzője, hogy egy speciális alkotóelemet; saját genetikai kóddal nem rendelkező, szelén tartalmú aminosavat (szelenociszteint) tartalmaznak. Mivel a szelenocisztein sokkal reaktívabb, mint a közönséges kén tartalmú cisztein – a szelenofehérjék reaktivitása is eltér a többi fehérjétől.
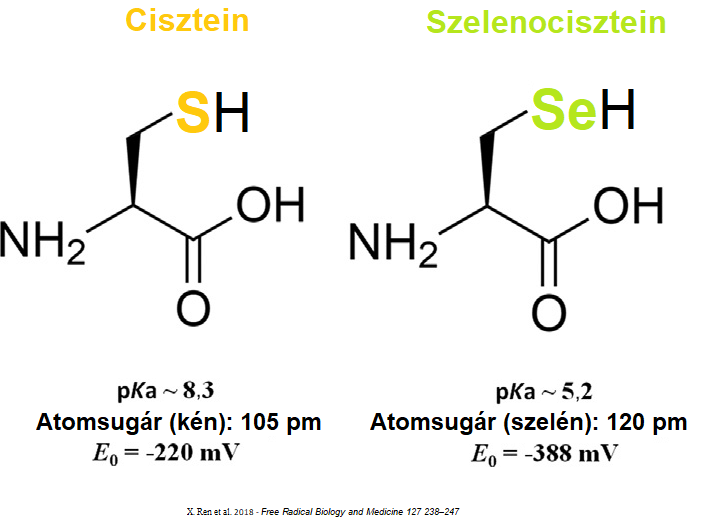
A szelenofehérjék jelenléte és működése létfontosságú az emberi szervezetben. Mára ismert, hogy szerepük van többek között a sejten belüli redox egyensúly szabályozásában, így a tumorok kialakulásában és a jelenleg használt kemoterápiás szerek hatásában is – ugyanakkor a pontos működési mechanizmusok és a kölcsönható partnerek még nem teljesen feltártak. Ezeknek az összefüggéseknek a felderítése és mélyebb megértése elengedhetetlen új terápiák fejlesztéséhez. Kutatási projektjeinkkel ennek a tudásbázisnak a bővítését célozzuk.
Laboratóriumunkban foglalkozunk szeléntartalmú és Trx-fold fehérjék rekombináns termelésével és in vitro karakterizálásával; a redox jelátviteli rendszer és transzkripciós válaszok vizsgálatával, továbbá potenciális daganatellenes szerek tesztelésével és tumoros betegek szérummintáinak elemzésére alkalmas, diagnosztikai célú ELISA-módszer beállításával is.
Eredményeink hozzájárulnak a rekombináns (laborkörülmények között előállított) szelenofehérjéken alapuló és a sejteken belüli redox szabályozás zavarait célzó új terápiák fejlesztéséhez.
Aktuális projektjeink
Rekombináns fehérje termelés: Szelenofehérjék, Trx-fold (tioredoxinhoz hasonló szerkezeti elemet tartalmazó fehérjék) és egyéb, redox-aktív fehérjék és ezek helyspecifikus mutánsainak előállítása E.coli-ban, tisztítása folyadékkromatográfiával, továbbá biokémiai jellemzése, összehasonlítása (pl. Trx1, Trx2, TXNL1, TXNRD1, TXNRD2, GPX3, PRX2). A létrehozott fehérjék alkalmasak tiszta enzimrendszereken alapuló vizsgálati rendszerek létrehozására (pl.gyógyszerjelölt molekulák azonosításához).

TXNL1 karakterizálása: A TXNL1 egy Trx-fold szerkezetű fehérje, melynek funkciója feltehetően a proteaszómához köthető. Ismert, hogy az Auranofin kezelés a TXNL1-szint gyors csökkenését váltja ki – ennek a hatásmechanizmusnak a felderítéséért az Auranofin celluláris hatását teszteljük TXNL1 knockdown és knockout humán sejtvonalakon. Célunk a TXNL1 fehérje még nem azonosított fiziológiás funkciójának felderítése az intracelluláris fehérje homeosztázisban.

pTRAF: A pTRAF módszer egy olyan plazmid-alapú technológia, amely lehetővé teszi humán sejtmodellekben egyszerre három jelátviteli útvonal aktiválódásának és egymásra hatásának vizsgálatát egy-sejt szinten. Az általunk létrehozott új pTRAF variánsok lehetővé teszik különböző, redox-szabályozott jelátviteli útvonalak feltárását, és azokra (és a tioredoxin rendszerre) specifikus inhibitorok vizsgálatát.

GPx izormák specifikus inhibitorainak vizsgálata: Laboratóriumunkban a National Institute of Health Transzlációs Kutatóközpontjában (NCATS, USA) nagy áteresztőképességű szűréssel azonosított GPx1- és GPx4-inhibitorok vizsgálatával foglalkozunk. Ellenőrizzük specificitásukat a tioredoxin család más fehérjéire, tiszta enzimrendszerekkel végzett vizsgálatokkal (Trx1, Trx2, TRP14, TXNL1). Emellett a szűrésből származó legígéretesebb gyógyszerjelöltek hatásait vizsgáljuk humán sejtvonalakon élősejtes képalkotó rendszerrel (Incucyte SX1).
Trx és GSH rendszerhez kapcsolódó biomarker kutatások: A Trx és GSH rendszerek redox biológiában betöltött szerepének tanulmányozása, valamint ezen rendszerek kulcsfontosságú fehérjéinek vizsgálata elsődleges célunk. Jelenleg ELISA-alapú módszerek kifejlesztése van folyamatban Trx1- és TrxR1-szint mérésére betegek szérummintáiban.




